DUPLICACIÓN.
Consiste en la formación de dos o mas cadenas de ADN a partir de una ya existente.
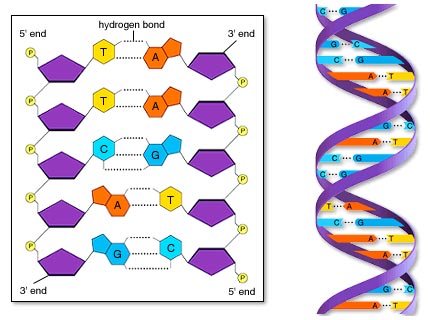
Esta duplicación se produce mediante un mecanismo semiconservativo, las dos cadenas complementarias del ADN original, al separarse, sirven de molde cada una para la síntesis de una nueva cadena complementaria de la cadena molde, de forma que cada nueva doble hélice contiene una de las cadenas del ADN original. Este experimento fue descubierto por Meselson y Sthal en 1958.
La duplicación es necesaria para la transmisión del material hereditario a las células hijas, esto solo se produce una vez y de forma completa en la vida de las células, en las células eucariotas se realiza en el periodo S del ciclo celular.
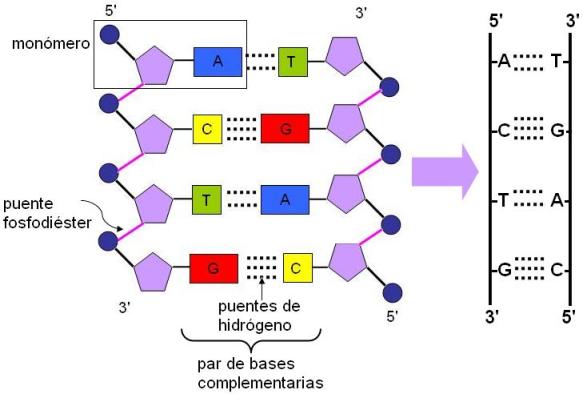
La molecula de ADN se abre como una cremallera por ruptura de los puentes de hidrógeno entre las bases complementarias.
Las enzimas que llevan a cabo la duplicación del ADN se llaman ADNpolimerasas y se han descubierto tres tipos: ADNpol I, ADNpol II y ADNpol III,
La ADNpol II es poco conocida aún y no se sabe su función específica, la ADNpol I interviene en la reparación del ADN y la ADNpol III es la que realmente intervienen en la duplicación de las cadenas del ADN.
Para que las polimerasas puedan funcionar se necesita que la doble hélice esté abierta, esto lo hacen un conjunto de proteínas.
Se abre la doble hélice mediante las helicasas.
Se estabiliza la apertura con las topoisomerasas y las proteínas SSB que impiden que las hebras separadas se superenrollen.
La ARNpolimerasa, sintetiza un cebador de ARN, empieza a sintetizarlo en alguna región rica en Adenina y timina.
La ADNpol III sintetiza la hebra complementaria del molde, uniendo nucleótidos al cebador. En un sentido de la horquilla de replicación la ADNpol III sintetiza la cadena de manera continua, pero en el sentido opuesto y debido a que las dos hebras del ADN original son antiparalelas, tiene que esperar a que se abra un poco la doble hélice y cuando reconoce un punto de inicio se forma el cebador y después un fragmento corto en sentido inverso a la hebra que se está sintetizando de manera continua, a medida que la horquilla se va abriendo se van sintetizando sucesivos fragmentos, llamados fragmentos de Okazaki, al final estos fragmentos se unen mediante la ADNligasa.
TRANSCRIPCIÓN.
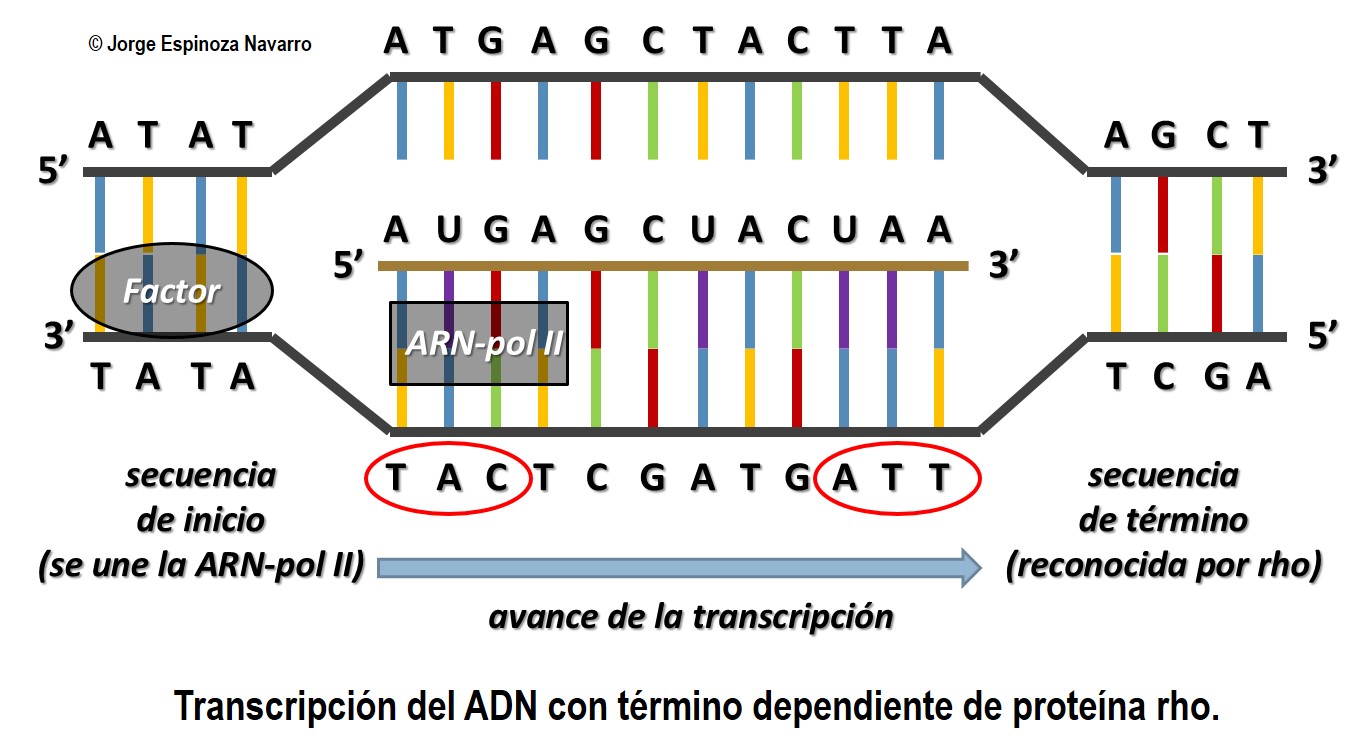
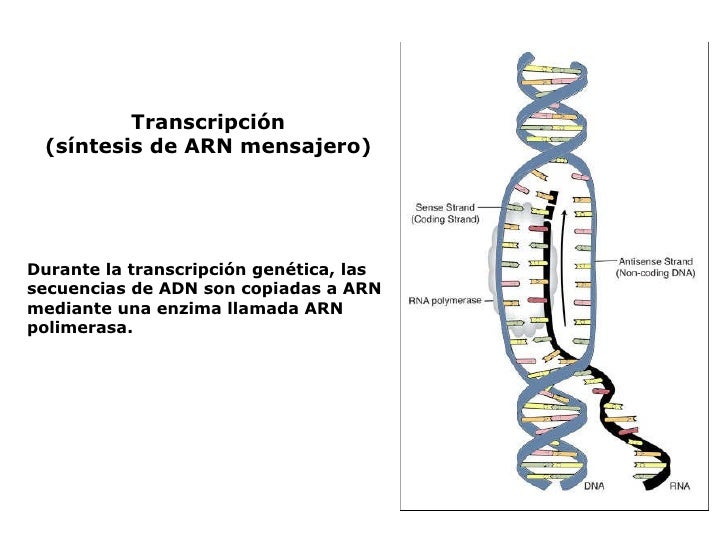
La TRANSCRIPCIÓN es el proceso de copia de un gen o fragmento de DNA utilizando
ribonucléotidos y originándose diferentes tipos de RNA.
Consiste en la formación de fragmentos de ARN utilizando como molde la cadena de ADN, estos fragmentos serán utilizados para ser leídos por los ribosomas y formar la proteína para la cual poseen la información (ARNm). También se han de transcribir, por supuesto, los genes del ARNr y los del ARNt.
Conviene recordar aquí que, lo que en cursos anteriores se llamaba función hereditaria de los ácidos nucleicos consistía en la división celular, pero además la función hereditaria consiste en controlar el metabolismo celular en cada momento de la vida de la célula, esto se realiza sintetizando de manera selectiva los ARNm de las proteínas que sean necesarias en cada momento. Por lo tanto y a diferencia de lo que ya hemos visto en la replicación, la transcripción es selectiva y reiterativa, se transcriben fragmentos concretos del ADN, no todo ello, y se pueden hacer más de una copia.
Solo se transcribe una hebra de las dos del ADN, siempre se hace de manera continua.
TRADUCCIÓN.
La traducción consiste en utilizar la información que está en forma de secuencia de nucleótidos para formar proteínas que son secuencias de aminoácidos.
Código genético:
El ARN es una secuencia de cuatro nucleótidos, las proteínas son secuencias de veinte aminoácidos.
El código genético es degenerado, hay 64 codones para solo 20 aminoácidos lo que significa que un aminoácido puede estar codificado por más de un codón, cuando ocurre esto el nucleótido que cambia en la combinación suele ser el último, siendo los dos primeros más estables.
Todas las proteínas empiezan a formarse por la combinación AUG, llamada codón de inicio, la lectura se hace sin saltar ningún nucleótido y sin que estos se utilicen en dos codones a la vez, el ribosoma "lee" sin interrupción desde el codón de inicio hasta que encuentra uno de los codones de terminación (UAA, UAG y UGA).
El código genético es universal, es el mismo para todos los seres vivos.
Ribosomas y traducción del material genético.
Son estructuras supramoleculares formadas por ARN y proteínas. Estas compuestos por dos subunidades.
En cuanto a la forma la subunidad menor, es alargada con una depresión en el tercio de su longitud, la subunidad mayor es semiesférica con tres protuberancias que forman una corona donde se encaja la subunidad menor. Se piensa que esta subunidad puede tener un canal por donde sale la proteína en formación.
Cuando hay varios ribosomas traduciendo la misma cadena de ARNm forman un polisoma.
Los ribosomas si están en el citoplasma forman proteínas que harán su función dentro de la célula, o pegados a la membrana del retículo endoplasmático, e este caso, las proteínas que formen podrán ser exportadas fuera de la célula.
Traducción del material genético. Síntesis de proteínas.
3 Fases:
Activación
El ARNm es una secuencia de nucleótidos que es reconocida por otra secuencia complementaria del ARNt, el ARNm no reconoce aminoácidos. Para que la traducción sea perfecta es necesario que la unión de un aminoácido a una cadena de proteínas, necesita dos pasos que han de estar controlados independientemente, la perfecta unión del aminoácido a su ARNt específico y posteriormente la correcta complementariedad del anticodón del ARNt con el codón del mensajero, si alguno de estos dos pasos falla puede producirse algún error en la traducción aunque el gen sea correcto.
La unión del aminoácido al ARNt es, por tanto, un mecanismo muy específico llevado a cabo por las enzimas AminoacilARNtsintetasas, hay una para cada ARNt. la reacción catalizada tiene dos fases; en la primera la enzima activa el aminoácido al unirlo a un nucleótido de adenina (AMP), procedente de la hidrólisis de ATP, el aminoácido se une por un enlace ester entre el carbono carboxílico del aminoácido y el fosfórico del nucleótido; posteriormente este enlace ester proporciona la energía para que el aminoácido se una al extremo CCA del ARNt, ahora la energía se encuentra en el enlace entre el grupo ácido del aminoácido y un grupo OH- de nucleótido de adenosina terminal del ARNt y será utilizado en la formación del enlace peptídico entre este aminoácido y el siguiente que se una a la cadena.
Iniciación
Comienza con la unión de la subunidad menor a la zona donde se encuentra el codón de iniciación AUG, el ribosoma discrimina entre un codón AUG de iniciación y otro en el interior de la secuencia gracias a otras secuencias de unión previas al AUG. AUG codifica para metionina que es el primer aminoácido que se coloca, el ARNt de iniciación y el que coloca la metionina en el interior de la cadena son diferentes.
Hay varios factores de iniciación IF (3 en procariotas y más de 3 en eucariotas) que en conjunto permiten la unión del ARNm con la subunidad menor, unen el ARNt de la metionina al mensajero y unen la subunidad mayor; en estos procesos se gasta una molécula de GTP.
El aminoácido queda instalado en un lugar físico de la subunidad mayor llamado centro peptidil.
Elongación
Es llevada a cabo por factores de elongación EFT y EFG. El EFT gastando una molécula de GTP acerca el siguiente ARNt y lo coloca enfrente de su codón, en el centro aminoacil de la subunidad mayor; el primer aminoácido que se encuentra en el centro peptidil se desprende de su ARNt y se une al grupo amino del recién colocado, la energía para formar el enlace peptídico procede de la hidrólisis del aminoácido de su ARNt (recordad fase de activación). El factor EFG gastando una molécula de GTP trasloca el dipéptido, unido al ARNt del último aminoácido y este unido al ARNm, del centro aminoacil al peptidil. Este proceso se repite para cada nuevo aminoácido, siempre se desprende el péptido del penúltimo ARNt para unirse al grupo amino del último que queda unido por su ARNt al ARNm y después se trasloca.
Para la colocación exacta de cada aminoácido se necesita una molécula de ATP en la activación y dos moléculas de GTP en la elongación, si sumamos además la energía de síntesis de cada aminoácido se puede considerar la síntesis de proteínas como un proceso energéticamente muy costoso, esta energía no queda almacenada como energía de enlace en la molécula, sino que ha sido empleada en el orden de los aminoácidos (aumento de entropía real aunque aparentemente ha aumentado el orden del sistema).
Terminación
Es producida por los factores de terminación R1 y R2 en presencia de los codones UAG,UGA y UAA. Los codones de terminación no son reconocidos por ningún ARNt pero sí por los factores de terminación que inducen la transferencia del péptido al agua, desprendiéndose del último ARNt al que está unido liberándose la proteína y separándose las dos subunidades del ribosoma.











 +
+ 

































{kind=link}
{kind=link}
{kind=link}